Вибір читачів

Популярні статті
Захворювання, одним із основних проявів яких є руйнування мієліну, – одна з найбільш актуальних проблем клінічної медицини, переважно неврології. Останніми роками спостерігається виразне збільшення кількості випадків захворювань, що супроводжуються ушкодженням мієліну.
Мієлін- особливий вид клітинної мембрани, що оточує відростки нервових клітин, в основному аксони, в центральній (ЦНС) та периферичній нервовій системі (ПНС).
Основні функції мієліну:
харчування аксона
ізоляція та прискорення проведення нервового імпульсу
опорна
бар'єрні функції.
За хімічним складом мієлін- це ліпопротеїдна мембрана, що складається з біомолекулярного ліпідного шару, розташованого між мономолекулярними шарами білків, що спірально закручена навколо інтернодального сегмента нервового волокна.
Ліпіди мієліну представлені фосфоліпідами, гліколіпідами та стероїдами. Всі ці ліпіди побудовані за єдиним планом і обов'язково мають гідрофобний компонент (хвіст) і гідрофільну групу (головку).
Білки становлять до 20% сухої маси мієліну. Вони бувають двох видів: білки, розташовані на поверхні, і білки, занурені в ліпідні шари або пронизують мембрану наскрізь. Усього описано понад 29 білків мієліну. Основний білок мієліну (ОБМ), протеоліпідний білок (ПЛП), мієлін-асоційований глікопротин (МАГ) становлять до 80% білка. Вони виконують структурну, стабілізуючу, транспортну функції, мають виражені імуногенні та енцефалітогенні властивості. Серед дрібних білків мієліну на особливу увагу заслуговує мієлін-олігодендроцитарний глікопротеїн (МОГ) і ферменти мієліну, що мають велике значення у підтримці структурно-функціональних взаємин у мієліну.
Мієліни ЦНС та ПНР відрізняються за своїм хімічним складом
в ПНЗ мієлін синтезується шванновськими клітинами, причому кілька клітин синтезують мієлін для одного аксона. Одна шванновская клітина утворює мієлін лише одного сегмента між ділянками без мієліну (перехопленнями Ранвье). Мієлін ПНР помітно товщі, ніж у ЦНС. Такий мієлін мають усі периферичні та черепні нерви, тільки короткі проксимальні сегменти черепних нервів та спинно-мозкових корінців містять мієлін ЦНС. Зоровий і нюховий нерви містять переважно центральний мієлін.
в ЦНС мієлін синтезується олігодендроцитами, причому одна клітина бере участь у мієлінізації кількох волокон.
Руйнування мієліну є універсальним механізмом реакції нервової тканини на ушкодження.
Хвороби мієліну поділяються на дві основні групи
мієлінопатії - пов'язані з біохімічним дефектом будови мієліну, як правило, генетично обумовленим
Мієлінокласті - в основі мієлінокластичних (або демієлінізуючих) захворювань лежить руйнування нормально синтезованого мієліну під впливом різних впливів, як зовнішніх, так і внутрішніх.
Підрозділ на ці дві групи вельми умовний, оскільки перші клінічні прояви мієлінопатій можуть бути пов'язані з впливом різних зовнішніх факторів, а мієлінокласті найімовірніше розвиваються у схильних осіб.
Найбільш поширене захворювання з усієї групи хвороб мієліну – розсіяний склероз. Саме з цим захворюванням доводиться найчастіше проводити диференціальну діагностику.
Спадкові мієлінопатії
Клінічні прояви більшості цих захворювань найчастіше відзначаються вже у дитячому віці. У той же час є ряд захворювань, які можуть починатися в пізнішому віці.
Адренолейкодистрофії (АЛД) пов'язані з недостатністю функції кори надниркових залоз і характеризуються активною дифузною демієлінізацією різних відділів як ЦНС, так і ПНР. Основний генетичний дефект при АЛД пов'язаний із локусом на Х-хромосомі - Xq28, генетичний продукт якого (білок ALD-P) є пероксисомальним мембранним білком. Тип успадкування у типових випадках – рецесивний, залежний від статі. В даний час описано більше 20 мутацій у різних локусах, пов'язаних з різними клінічними варіантами АЛД.
Основний метаболічний дефект при цьому захворюванні - збільшення вмісту в тканинах насичених жирних кислот з довгим ланцюгом (особливо С-26)що призводить до грубих порушень структури та функцій мієліну. Поряд із дегенеративним процесом у патогенезі хвороби істотне значення має хронічне запалення у тканині мозку, пов'язане з підвищеною продукцією фактора некрозу пухлин альфа (ФНП-a). Фенотип АЛД визначається активністю цього запального процесу і найімовірніше обумовлений як різним набором мутацій на Х-хромосомі, і аутосомной модифікацією впливу дефектного генетичного продукту, тобто. поєднанням основного генетичного дефекту в статевому Х-хромосомі зі своєрідним набором генів на інших хромосомах.
За типом успадкування та особливостями початку АЛД класифікують на
Зчеплені з Х-хромосомою АЛД у дітей
зчеплені з Х-хромосомою АЛД у дорослих
аутосомнорецесивні неонатальні АЛД.
Клінічні варіанти адренолейкодистрофії можна розділити на дві основні групи
Переважно церебральні форми
переважно поліневропатичні, що частіше розвиваються у дорослих.
Залежно від переважаючого симптомокомплексу у складі АЛД крім поліневритичного виділяють кілька клінічних варіантів:
1) прогресуюча енцефалопатія з кірковою сліпотою у молодих людей;
2) прогресуюча мієлопатія у зрілих чоловіків;
3) енцефалопатія у поєднанні з мієлопатією;
4) злоякісна енцефалопатія у новонароджених хлопчиків;
5) переважно ендокринологічні порушення (хвороба Аддісона), зчеплені з Х-хромосомою та супроводжуються помірними неврологічними порушеннями;
6) мієлопатія у жінок - носіїв патологічного гена та ін.
У типових випадках на церебральні форми АЛД страждають хлопчики 3 - 15 років, відзначаються прогресуючі порушення рухів, зниження зору, слуху, типові судомні синдроми та деменція. Окремі симптоми енцефалопатії та мієлопатії можуть бути виявлені і у гетерозиготних жінок – носіїв патологічного гена.
Серед рідкісних поліневритичних форм виділяютьадреномієлоневропатію, яка може починатися у віці 25 - 35 років і характеризується прогресуючою мієлопатією та поліневропатією. Клінічні прояви: наростаюча слабкість у ногах, порушення чутливості за поліневритичним типом ("шкарпетки" та "рукавички"), порушення координації. Для проведення диференціальної діагностики велике значення має сімейний анамнез, що неухильно прогресує тип перебігу захворювання, зниження в крові вмісту адреналіну та збільшення довголанцюгових жирних кислот.
Останнім часом дедалі більшу роль моніторингу активності патологічного процесу при різних варіантах АЛД набувають магнітно-резонансна томографія.
Ефективного специфічного лікування АЛД поки що немає, проводиться симптоматична терапія Наразі розробляються підходи до проведення генетичного консультування як методу профілактики АЛД на основі скринінгу мутацій у гені білка ALD та інших локусах для виявлення носіїв патогенних мутацій.
Описано пізню форму суданофільної лейкодистрофії Пеліцеуса-Мерцбахера з початком захворювання на другому десятилітті життя. Виражене демієлінізуюче ураження головного мозку у цих хворих супроводжується зниженням вмісту ефірів холестерину. У цих хворих прогресивно зростають порушення координації, спастичні парези, інтелектуальні порушення.
Пізні форми цієї лейкодистрофії часто мають сприятливий перебіг, і їх, як правило, дуже складно диференціювати від розсіяного склерозу.
Хвороба Александера рідкісне захворювання, переважно успадковане за аутосомно-рецесивним типом, входить до групи лейкодистрофій, які характеризуються демієлінізацією з дифузною волокнистою дегенерацією білої речовини головного мозку та утворенням у тканині мозку глобоїдних клітин.
Ця дисмієлінопатія характеризується тим, що в мієлін замість галактоліпідів і цереброзидів накопичуються глюколіпіди.
Для неї характерні
спастичні паралічі, що поступово наростають
зниження гостроти зору
деменція
епілептичний синдром
гідроцефалія.
У тканині мозку при хворобі Александера, крім дифузної демієлінізації, виявляються порожнини та гіалінозні ущільнення волокон навколо судин мозку.
Захворювання може починатися в будь-якому віці, на ранніх стадіях можливе ремітування.
Прогноз вкрай несприятливий, хоч і описані повільно прогресуючі варіанти, близькі за течією до розсіяного склерозу.
До групи глобоїдо-клітинних лейкодистрофій також належать такі рідкісні захворювання, як хвороба Краббе та хвороба Канавана
. Ці захворювання рідко розвиваються у дорослому віці.
Клінічно вони характеризуються прогресуючим поразкою мієліну різних відділів ЦНС з недостатнім розвитком парезів, порушень координації, деменції, сліпоти, епілептичним синдромом. Дослідження лейкоцитів крові або фібробластів шкіри часто виявляють дефіцит ферменту b-галактоцереброзідази.
У різному віці можуть починатися так звані хвороби мітохондрій із поразкою ЦНС та розвитком мітохондріальних лейкоенцефалопатій . Різні мутації, у тому числі в мітохондріальній ДНК, можуть викликати ферментативні дефекти та порушувати процеси транспорту та утилізації речовин, збереження та передачі енергії, порушувати дихальний ланцюг.
Для мітохондріальних енцефалопатій характерно про підвищення вмісту лактату та пірувату у плазмі крові та спинно-мозкової рідини, у мітохондріях. Ці порушення можуть призводити до розвитку поліморфної неврологічної симптоматики, іноді подібної до симптомів розсіяного склерозу. Часто мітохондріальна енцефалопатія проявляється слабкістю та швидкою стомлюваністю м'язів, офтальмоплегією та іншими симптомами.
Мієлінокластичні захворювання
Серед мієлінокластичних захворювань на особливу увагу заслуговують вірусні інфекції, у патогенезі яких велике значення має руйнування мієліну. Це насамперед
нейроСНІД, що викликається вірусом імунодефіциту людини (ВІЛ), та пов'язані з ним ураження нервової системи
тропічний спінальний парапарез (ТСП)
, що викликається ретровірусом HTLV-I
Патогенез первинного ураження ЦНС при нейроСНІД пов'язаний з безпосереднім нейротоксичним впливом вірусу, а також з патологічною дією цитотоксичних Т-клітин, протимозкових антитіл та нейротоксичних речовин, що виробляються інфікованими імуноцитами. Пряме ураження мозку при ВІЛ-інфекції призводить до розвитку підгострого енцефаліту з ділянками демієлінізації. При цьому в тканині мозку можуть бути виявлені моноцити та макрофаги з великою кількістю вірусу. Ці клітини зливаються в гігантські багатоядерні утворення з величезною кількістю вірусного матеріалу (синцитії), що і стало причиною позначення цього енцефаліту як гігантоклітинного. У той же час характерною для цієї інфекції є невідповідність тяжкості клінічних проявів та виразності патоморфологічних змін. У багатьох хворих із виразними клінічними проявами ВІЛ-асоційованої деменції патоморфологічно може виявлятися лише "збліднення" мієліну та слабовиражений центральний астрогліоз.
Клінічно найчастіше при ВІЛ-інфекції відзначається так званий ВІЛ-асоційований пізнавально-руховий комплекс.
У цей комплекс, що позначався раніше як СНІД-деменція, включають три захворювання:
ВІЛ-асоційовану деменцію
ВІЛ-асоційовану мієлопатію
ВІЛ-асоційовані мінімальні пізнавально-рухові розлади.
При найчастішому синдромі, ВІЛ-деменції, виявляється уповільнення психомоторних процесів, неуважність, зниження пам'яті. Надалі деменція швидко прогресує, можливе приєднання осередкових неврологічних симптомів. При ВІЛ-асоційованій вакуольні мієлопатії переважають рухові розлади, переважно в нижніх кінцівках, пов'язані з ураженням спинного мозку.
У хворих із імунодефіцитами різного генезу може розвиватися прогресуюча мультифокальна лейкоенцефалопатія (ПМЛ) яка викликається поліомавірусом (вірус JC з групи паповавірусом). Крім хворих на СНІД, ця інфекція може мати місце у хворих з саркоїдозом, туберкульозом, ВКВ, лімфомою, лейкемією, при імунодефіцитах після пересадки нирки. Вірус виявляється в олігодендроцитах та астроцитах, причому останні можуть зливатися, утворюючи гігантські клітини. У патогенезі демієлінізації має значення як безпосереднє ураження олігодендроцитів, так і аутоімунні реакції на фоні імунодефіциту. Для ПМЛ характерний неухильно прогресуючий перебіг із раннім розвитком деменції, екстрапірамідними порушеннями, дезорієнтацією в місці та часі. Вирішальне значення у діагностиці ПМЛ має виявлення у клітинах спинно-мозкової рідини JC-вірусу.
Хронічне запалення в білій речовині мозку з розвитком демієлінізації характерно для ТСП (тропічного спінального парапарезу) або HTLV-1 мієлопатії . Це ендемічне для країн Південно-Східної Азії захворювання характеризується неухильно прогресуючим нижнім парапарезом. При проведенні МРТ у хворих з ТСП можуть виявлятися субклінічні осередки демієлінізації в головному мозку. Діагностичне значення має виявлення ретровіруса HTLV-I чи антитіл щодо нього.
Лікування всіх вірусних інфекцій ґрунтується на використанні противірусних препаратів, що зупиняють розмноження вірусу в інфікованих клітинах.
У осіб з кахексією, які страждають на хронічний алкоголізм, тяжкі хронічні захворювання печінки та нирок, при діабетичному кетоацидозі, при проведенні реанімаційних заходів може розвиватися тяжке демієлінізуюче захворювання. гострий або підгострий центральний понтійний та/або екстрапонтійний мієліноліз.
При цьому захворюванні симетричні білатеральні вогнища демієлінізації утворюються у підкіркових вузлах та стовбурі головного мозку. Передбачається, що основою цього процесу є порушення у балансі електролітів, насамперед іонів Na.
Особливо високий ризик розвитку мієлінолізу при швидкій корекції гіпонатріємії.
Клінічноцей синдром може проявлятися як мінімальними неврологічними симптомами, так і важкими альтернуючими синдромами та розвитком коми. Захворювання зазвичай через кілька тижнів закінчується смертю, але в ряді випадків масивні дози кортикостероїдів запобігають летальному исходу.
Після опромінення, хіміо- та радіотерапії може розвиватися токсична лейкоенцефалопатія
з осередковою демієлінізацією у поєднанні з мультифокальним некрозом. Можливий розвиток
гострих
ранніх відстрочених
пізніх демієлінізуючих процесів.
Останні починаються через кілька місяців або років після опромінення та характеризуються тяжким перебігом з поліморфною осередковою неврологічною симптоматикою. У патогенезі цих лейкоенцефалопатій важливе значення мають аутоімунні реакції на антигени мієліну, ушкодження олігодендроцитів і, отже, порушення ремієлінізації.
Токсичне пошкодження мієліну може спостерігатися також при
порфірії
гіпотиреозі
інтоксикація ртуттю
свинцем
СО
ціанідами
при всіх видах кахексії
передозування антиконвульсантів
ізоніазиду
актиноміцину
при вживанні героїну та морфію
На особливу увагу заслуговує ряд мієлінокластичних захворювань, які можуть бути класифіковані як особливі варіанти розсіяного склерозу.
Початок розсіяного склерозу клінічно дуже складно відрізнити від гострого розсіяного енцефаломієліту (ОРЕМ)
- аутоімунного запального захворювання ЦНС, що розвивається, як правило, після інфекцій або вакцинацій. Диференціювати від розсіяного склерозу у багатьох випадках можна лише на основі даних динамічного спостереження
при ОРЕМ процес завжди однофазний, тобто. гострий
при РС – хронічний.
В більшості випадків ОРЕМ розвивається після інфекційного захворювання чи вакцинації.
Серед поствакцинальних енцефалітів особливо часто описують енцефаліти після введення вакцин проти кашлюку та віспи, а також після антирабічних щеплень.
ОРЕМ може розвиватися після або під час
вірусних захворювань (кору, краснухи, віспи, вітряної віспи, епідемічного паротиту, інфекційного мононуклеозу, герпетичної інфекції, грипу)
рідше - при мікоплазмовій
іншої бактеріальної інфекції.
Патогенез захворювання заснований на розвитку аутоімунної реакції та гострого запалення через перехресні реакції між антигенами вірусів (рідше бактерій) та антигенами мозку.
У типових випадках є латентний період 4 - 6 днів. Захворювання починається гостро або підгостро, із загальноінфекційної, загальномозкової та осередкової симптоматики, у важких випадках – із судомним синдромом та порушенням свідомості.
Часто ОРЕМ поєднується з гострим поліневритом на кшталт моторного поліневриту Гійєна - Барре.
На МРТ при ОРЭМ з'являються множинні осередки підвищеної інтенсивності на Т2-зважених зображеннях, що активно накопичують контраст у Т1-режимі. Ці вогнища дуже близькі до спостережуваних при розсіяному склерозі, але при ОРЕМ вони часто більші за розміром, іноді симетричні, із залученням таламуса та базальних ядер. При ОРЕМ на повторних томограмах не відзначається появи нових вогнищ, як при розсіяному склерозі, а навпаки, ряд вогнищ може зменшуватися у розмірах.
Лікування ОРЕМ засноване на використанні пульс-доз кортикостероїдів та адекватної симптоматичної терапії. У ряді випадків зареєстрований добрий клінічний ефект від використання плазмоферезу, великих доз імуноглобуліну класу G (IgG) внутрішньовенно.
(!!!) Злоякісні форми ОРЕМ з розвитком множинних геморагій у тканину мозку іноді класифікують якгострі геморагічні лейкоенцефаліти чи хвороба Херста.
Особливий варіант розсіяного склерозу був описаний Девісом у 1894 р. та отримав назву оптикомієліт Девіка . Для цього захворювання характерні симптоми ураження спинного мозку (поперечний мієліт) і зорових нервів з однієї або двох сторін, а також злоякісний неухильно прогресуючий перебіг та поганий прогноз. Найчастіше випадки хвороби Девіса трапляються у жінок корінних національностей країн Південно-Східної Азії. Патоморфологіческіе вогнища демієлінізації при цьому захворюванні близькі до розсіяного склерозу, але іноді спостерігаються нетипові для останнього дифузні запальні зміни з вираженим набряком тканини мозку і в окремих випадках з геморагіями. Дані МРТ та аутопсії свідчать про можливість утворення бляшок не тільки в спинному мозку та зорових нервах, а й у перивентрикулярній білій речовині, рідко у стовбурі мозку та мозочку. У патогенезі оптикомієліту Девіса велике значення мають гуморальні аутоімунні реакції з утворенням антитіл до різних антигенів нервової тканини.
Концентричний склероз, чи хвороба Балло , є неухильно прогресуючим демієлінізуючим захворюванням осіб молодого віку При цьому захворюванні утворюються великі осередки демієлінізації переважно в білій речовині лобових часток, іноді із залученням сірої речовини. Вогнища складаються з областей повної і часткової демієлінізації з вираженим раннім ураженням олігодендроцитів. Зони демієлінізації та ремієлінізації при цьому розташовані концентрично або хаотично; це надає вогнищам на томограмах характерний вигляд. До застосування МРТ діагноз хвороби Балло підтверджувався лише за аутопсии. Прогноз украй несприятливий. В останні роки отримано дані про поліпшення у стані хворих на тлі активної імуносупресії, що робить прогноз менш фатальним, як раніше.
Лейкоенцефаліт Шильдера частіше хворіють діти, хоча нерідко це захворювання починається у дорослому віці. Для нього характерна поширена неухильно прогресуюча демієлінізація з утворенням зливних зон запалення та руйнування мієліну. Типово поширення такої зони демієлінізації з однієї півкулі на іншу через мозолисте тіло. Клінічна картина хвороби Шільдера залежить від обсягу та локалізації ураження.
Характерно розвиток
епілептичного синдрому,
психозів,
центральних парезів,
гіперкінезів, атаксії,
зниження зору,
деменції,
псевдобульбарного синдрому
Іншим злоякісним варіантом розсіяного склерозу є хвороба Марбурга , описана як прогресуюче захворювання з гострим початком, переважним ураженням стовбура мозку, вкрай злоякісним, з швидко прогресуючим перебігом та швидким смертельним наслідком. Крім стовбура мозку, множинні осередки демієлінізації при цьому захворюванні часто локалізуються в зорових нервах та шийному відділі спинного мозку. Патоморфологічні осередки демієлінізації при хворобі Марбурга характеризуються швидким розвитком дегенерації аксонів, іноді некрозом. Активна імуносупресія у ряді випадків сприяла настанню ремісії, що підтверджує спільність хвороби Марбурга та розсіяного склерозу.
(!!!) Слід зазначити, що осередки демієлінізації в ЦНС досить часто виявляються у хворих з
-системним червоним вовчаком
-первинним синдромом Шегрена
-васкулітами різного генезу та іншими системними аутоімунними захворюваннями
Руйнування мієліну та розвиток аутоімунних реакцій на його компоненти спостерігається при багатьох судинних та паранеопластичних процесах у ЦНС, що слід враховувати під час проведення диференціального діагнозу.
,
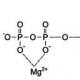
Основний білок мієліну(також основний білок мієліну, англ. myelin basic protein) - являє собою основний (порядку 50%) білковий компонент мієліну. При електрофоретичному аналізі в ПААГ з екстрактами SDS мієлінової оболонки людини визначається до 10 смуг. Основний білок мієліну знаходиться в комплексі з ліпідами та має молекулярну масу близько 30 кДа.
Для основного білка мієліну людини існує поліморфізм послідовностей амінокислот у 46 та 47 положенні. У цьому положенні знаходяться залишки гліцину, часто у поєднанні залишками серину. Було показано, що у хворих на розсіяний склероз відзначалася заміна серину на гліцин у 44-49 положеннях. Поруч авторів було показано високий рівень гомології - близько 80-90 % амінокислотної послідовності основного білка мієліну в різних видів тварин. Так, амінокислотні послідовності основного білка мієліну людини і бика відрізняються лише по залишках амінокислот в кількох положеннях, в той час як основний білок мієліну щура відрізняється від основного білка мієліну людини і бика положенням 40 амінокислотних залишків в середині полипептид7 ланцюга (від 1 .
Основний білок мієліну містить значний відсоток (приблизно 25 %) основних амінокислот (аргінін, лізин та гістидин), рівномірно розподілених по всьому поліпептидному ланцюгу, що зумовлює дуже високу ізоелектричну точку (рI = 12-13). Такі особливості призводять до взаємодії всієї молекули основного білка мієліну з карбоксильними групами мембранних ліпідів. Основний білок мієліну є мембранним білком і має значну спорідненість до ліпідів, в комплексі з якими виділяється. Основний білок мієліну є полікатіоном і формує особливо стабільні комплекси з аніонними ліпідами. Незважаючи на те, що ступінь зв'язування основного білка мієліну значно вищий з кислими ліпідами, він досить виражений і щодо цвіттер-іонів, особливо щодо сфінгомієліну. Основний білок мієліну також взаємодіє з фосфатидилетаноламіном при рН 7,2, коли цей ліпід знаходиться переважно в цвіттер-формі. Незвичайна здатність основного білка мієліну утворювати нерозчинні комплекси з додецилсульфатом натрію при значеннях рН вище та нижче ізоелектричної точки. При формуванні комплексів основного білка мієліну з основною масою аніонних фосфоліпідів, однаково відіграють роль як гідрофобні, так і електростатичні взаємодії. Найбільш виражений вплив на стабілізацію ліпідів основний білок мієліну вказує при вбудовуванні в ліпідні хвости. p align="justify"> Методом рентгеноструктурного аналізу було показано, що при формуванні мієліну відбувається занурення більшої частини молекул основного білка мієліну в шар головок ліпідів.
Після введення основного білка мієліну в ЦНС розвиваються симптоми, схожі на симптоми розсіяного склерозу. Поява цих симптомів отримала назву експериментального алергічного енцефаломієліту.
Мієлінова оболонка нервів на 70-75% складається з ліпідів і на 25-30% - з білків. До складу її клітин також входить лецитин – представник фосфоліпідів, роль якого дуже велика: він бере участь у багатьох біохімічних процесах, покращує опірність організму до токсинів, знижує рівень холестерину.
Вживання продуктів, що містять лецитин, є гарною профілактикою та одним із способів лікування захворювань, пов'язаних із порушенням діяльності нервової системи. Ця речовина входить до складу багатьох круп, сої, риби, яєчного жовтка, пивних дріжджів. Лецитин також містять: печінку, оливки, шоколад, родзинки, насіння, горіхи, ікра, молочні та кисломолочні продукти. Додатковим джерелом цієї речовини можуть бути біологічно активні харчові добавки.
Відновити мієлінову оболонку нервів можна, включаючи дієту продукти, що містять амінокислоту холін: яйця, бобові, яловичина, горіхи. Дуже корисні омега-3 поліненасичені жирні кислоти. Вони містяться в рибі жирних сортів, морепродуктах, насінні, горіхах, лляному маслі та лляному насінні. Джерелом омега-3 жирних кислот можуть бути: риб'ячий жир, авокадо, волоські горіхи, квасоля.
До складу мієлінової оболонки входять вітаміни В1 і В12, тому для нервової системи буде корисно включати до раціону житній хліб, цільнозернові крупи, молочні продукти, свинину, свіжу зелень. Дуже важливо вживати достатню кількість фолієвої кислоти. Її джерела: бобові (горох, квасоля, сочевиця), цитрусові, горіхи та насіння, спаржа, селера, броколі, буряк, морква, гарбуз.
Відновленню мієлінової оболонки нервів сприяє мідь. Її містять: кунжут, гарбузове насіння, мигдаль, гіркий шоколад, какао, свиняча печінка, морепродукти. Для здоров'я нервової системи необхідно включати до раціону продукти, що містять інозитол: овочі, горіхи, банани.
Дуже важливо підтримувати імунну систему. За наявності в організмі джерел хронічного запалення або автоімунних захворювань порушується цілісність нервів. У цих випадках, на додаток до основної терапії потрібно вводити в меню харчові та трав'яні протизапальні засоби: зелений чай, настої шипшини, кропиви, деревію, а також продукти, багаті на вітаміни С і Д. Вітамін С у великій кількості міститься в цитрусових, ягодах, ківі, капусті, солодкому перці, томатах, шпинаті. Джерелами вітаміну Д є яйця, молочні продукти, вершкове масло, морепродукти, жирні сорти риби, печінку тріски та інших риб.
Дієта для відновлення мієлінової оболонки нервів має містити достатню кількість кальцію. Він входить до складу багатьох продуктів: молоко, сир, горіхи, риба, овочі, фрукти, крупи. Для повноцінного засвоєння кальцію необхідно включати до раціону магній (міститься в горіхах, хлібі грубого помелу) і фосфор (міститься в рибі).
1Проведено вивчення концентрації основного білка мієліну (ОБМ) та нейронспецифічної енолази (НСЕ) у сироватці крові у 84 хворих на хронічний гепатит (ХГ) (вірусної етіології HBV, HCV – 38; алкогольної етіології – 17; аутоімунний гепатит – 11; ) та 77 цирозом печінки (ЦП) (вірусної етіології HBV, HCV, HBV+HCV – 27; первинний біліарний цироз – 10, алкогольної етіології – 18; змішаної етіології – 22). Контрольна група – 30 практично здорових осіб (донори). Концентрації ОБМ та НСЕ у сироватці крові визначали методом твердофазного імуноферментного аналізу з використанням комерційних тест-систем 449-5830 DSL МВР та 420-10 Fujirebio НСЕ. За результатами проведеного дослідження при алкогольних ураженнях печінки як на стадії ХГ, так і ЦП, що сформувався, спостерігалося значне підвищення концентрації ОБМ крові в порівнянні з вірусними ураженнями. Концентрація НСЕ у хворих на ЦП досліджуваних етіологічних груп, на відміну від ХГ, суттєво не відрізнялася.
основний білок мієліну
нейронспецифічна енолаза
хронічний гепатит
цироз печінки
печінкова енцефалопатія.
1. Жукова І.А. Нейронспецифічна енолаза як неспецифічний маркер нейродегенеративного процесу/І.А. Жукова, В.М. Аліфірова, Н.Г. Жукова// Бюлетень сибірської медицини. - 2011. - Т. 10. - № 2. - С. 15-21.
2. Білопасов В.В. Клінічна диференціація печінкової енцефалопатії у хворих на цироз печінки / В.В. Білопасов, Р.І. Мухамедянова, М.К. Андрєєв, Б.М. Левітан // Вятський медичний вісник. – 2002. – № 1. – С. 46-47.
3. Івашкін В.Т. Захворювання печінки та печінкова енцефалопатія / В.Т. Івашкін, Ф.І. Комаров, І.О. Іваніков // Російський медичний журнал. - 2001. - Т. 3. - № 12. - С. 150-155.
4. Левітан Б.М. Хронічна патологія печінки та кишковий мікробіоценоз (клініко-патогенетичні аспекти) / Б.М. Левітан, А.Р. Умерова, Н.М. Ларіна. - Астрахань: АГМА, 2010. - 135 с.
5. Левітан Б.М. Зміни концентрації основного білка мієліну у сироватці крові при захворюваннях печінки / Б.М. Левітан, А.В. Астахін, О.О. Євлашева // Експериментальна та клінічна гастроентерологія. - 2015. - № 2. - С. 93.
6. Павлов Ч.С. Печінкова енцефалопатія: патогенез, клініка, діагностика, терапія/Ч.С. Павлов, І.В. Дамулін, В.Т. Івашкін // Російський журнал гастроентерології, гепатології, колопроктології. - 2016. - № 1. - С. 44-53.
7. Торопова Н.Є. Оцінка інформативності нейронспецифічної енолази, що визначається імуноферментним методом / Н.Є. Торопова, Є.А. Дорофєєва, С.П. Дворянінова, Ж.П. Васієва // Клінічна лабораторна діагностика. – 1995. – № 1. – С. 15–17.
8. Чехонін В.П. Основний білок мієліну. Будова, властивості, функції, що у діагностиці деміелінізуючих захворювань / В.П. Чехонін, О.І. Гуріна, Т.Б. Дмитрієва та співавт. // Біомедична хімія. - 2000. - Т. 46. - № 6. - С. 549-563.
9. Arguedas M.R. Influence of hepatic encephalopathy on healthy – пов'язана з якістю життя в пацієнтів з cirrhosis / M.G. Arguedas, T.G. Delawrence, В.М. Mcguire // Digestive diseases and sciences. - 2003. - V. 48. - P. 1622-1626.
10. Butterworth R.F. Pathophysiology of hepatic encephalopathy: The concept of synergism // Hepatol. Res. – 2008. – V. 38. – P. 116-121.
11. Isgro M. Neuron – специфічний enolase як biomarker: biochemical and clinic aspects / M. Isgro, P. Bottoni, R. Scatena // AdvExp Mad Biol. - 2015. - Vol. 867. - P. 125-143.
12. Persson L. 100 proteínів і нейрон-специфічних ендола в cerebrospinal fluid and serum: маркери глобулю в людській центральній нервовій системі / L. Persson, H.G. Hardemark, J. Gustaffson та ін. // Stroke. – 1987. – Vol. 18. – P. 911-918.
13. Rabinowicz A. NSE є useful prognostic factor для пацієнтів після cerebral hypoxia / A. Rabinowicz, H. Reiber // Epilepsia. – 1996. – Vol. 37. – P. 122-125.
14. Tzakos A. Структура і функція м'язових proteínів: поточний статуй і виразки в відношенні до multiple sclerosis / A. Tzakos, A. Troganis, V. Theodorou // Curr. Med. Chem. - 2005. - Vol. 12. - P. 1569-1587.
Хронічний гепатит (ХГ) та цироз печінки (ЦП) є поліетиологічними захворюваннями. Загальновідомо, що інфікування гепатотропними вірусами є основним етіологічним чинником, що призводить до розвитку ХГ, а зловживання алкоголем, своєю чергою, є другою основною причиною цієї патології.
Перебіг та прогноз захворювань печінки значною мірою визначається наявністю та ступенем вираженості ураження центральної нервової системи (ЦНС). Печінкова енцефалопатія (ПЕ) є комплексом потенційно оборотних нервово-психічних порушень, обумовлених ураженням ЦНС токсичними речовинами, не знешкодженими патологічно зміненою печінкою, що виникають насамперед у результаті гострої або хронічної печінкової недостатності. Враховуючи крайню агресивність цих речовин, можна припустити, що під їх впливом відбувається деструкція нервової тканини з вивільненням рідких середовищ організму продуктів її розпаду.
Вивченню діагностичної та прогностичної значущості таких маркерів нейродеструкції, як основний білок мієліну (ОБМ) та нейронспецифічна енолаза (НСЕ), при різних патологічних станах ЦНС присвячено досить велику кількість досліджень. У той самий час питання їх діагностичної цінності при хронічних дифузних захворюваннях печінки (ХДЗП) різної етіології залишається мало вивченим. У зв'язку з цим вивчення ОБМ та НСЕ залежно від етіології ХДЗП є актуальним та перспективним.
Мета: вивчити діагностичну значущість визначення концентрації основного білка мієліну та нейронспецифічної енолази у сироватці крові залежно від етіології ХДЗП.
Матеріали та методи. Для вирішення поставлених завдань за період з 2012 по 2014 р. було обстежено 84 хворих на ХГ (вірусної етіології HBV, HCV – 38; алкогольної етіології – 17; аутоімунний гепатит – 11; змішаної етіології – 18) та 77 ЦП (вірусної етіології HV , HBV+HCV - 27;первинний біліарний цироз - 10, алкогольної етіології - 18;змішаної етіології - 22), які перебували на стаціонарному лікуванні в гастроентерологічному відділенні ГБУЗ АТ «АМОКБ». Серед обстежених хворих з патологією печінки було виділено групу з 17 пацієнтів, яка не увійшла до складу хворих на ХГ. Цю групу становили пацієнти з гострим алкогольним гепатитом (ОАГ), що протікає з симптомами вираженої печінково-клітинної недостатності. Контрольну групу становили 30 практичних здорових осіб (донори).
Дослідження проводилися на основі власних спостережень та даних медичної документації (клінічна історія хвороби, амбулаторна карта, висновки фахівців із параклінічним методам обстеження).
Хворі надходили до клініки на стадії загострення основного захворювання. При постановці діагнозу використані загальноприйняті класифікації. Клінічний діагноз встановлювався на підставі скарг хворих, вивчення анамнезу, фізикальних даних, лабораторних та інструментальних методів дослідження. В анамнезі приділяли особливу увагу оперативним втручанням, гемотрансфузіям, вживанню алкоголю та внутрішньовенних наркотичних препаратів, тривалому застосуванню гепатотоксичних лікарських препаратів, наявності спадкових захворювань.
Критерії виключення: супутня патологія ЦНС, лікування препаратами, що мають побічний нейротоксичний ефект.
Концентрації ОБМ та НСЕ у сироватці крові визначали методом твердофазного імуноферментного аналізу за допомогою наборів реагентів комерційних тест-систем 449-5830 DSL МВР та 420-10 Fujirebio НСЕ.
Статистичну обробку даних виконували з використанням пакета прикладних програм Statistica 6.0. Для кількісного порівняння ознак двох незв'язаних груп застосовували параметричний критерій Стьюдента (t). Кореляційний аналіз із розрахунком коефіцієнта кореляції (r) проводили з використанням критерію Спірмена. Відмінності вважали статистично значущими при досягнутому рівні значимості p<0,05.
Результати та обговорення. Концентрація ОБМ у хворих на ХГ вірусної етіології в середньому склала 1,9±0,27 нг/мл, змішаної - 2,3±0,3 нг/мл, аутоімунної 2,17±0,19 нг/мл, що достовірно не відрізнялося від результатів, отриманих у групі донорів – 1,9±0,3 нг/мл (р>0,05) (рис. 1). Більш значне підвищення рівня ОБМ було виявлено у хворих на хронічний гепатит алкогольної етіології, склавши 2,9±0,39 нг/мл, що достовірно перевищувало значення, отримані в контрольній групі, а також у хворих на вірусну етіологію захворювання (р<0,05). Максимальная концентрация ОБМ в сыворотке крови была выявлена в группе больных ОАГ, составив в среднем 5,4±0,17 нг/мл, что достоверно превышало показатели, характерные как для здоровых лиц, так и для больных хроническим гепатитом вирусной, смешанной, аутоиммунной и алкогольной этиологии (р<0,05). В исследуемой группе пациентов ОАГ максимальная концентрация ОБМ в периферической крови наблюдалась в 75% случаев.
Результати, отримані при дослідженні концентрації НСЕ у хворих на ХГ та ОАГ, дещо відрізнялися (рис. 2).
Концентрація НСЕ у хворих на хронічний гепатит вірусної етіології склала 6,9±0,41 нг/мл, змішаної - 7,4±0,37 нг/мл, аутоімунної - 6,4±0,52 нг/мл. Отримані результати близькі і достовірно не відрізнялися від значень, одержаних у контрольній групі - 6,49±0,41 нг/мл (р>0,05).
Рівень НСЕ у хворих на хронічний гепатит алкогольної етіології в середньому склав 8,1±0,51 нг/мл, що достовірно вище, ніж у контрольній групі, а також у хворих на аутоімунний та ХГ вірусної етіології (р<0,05).
Найбільш значне підвищення концентрації НСЕ, як і ОБМ, було виявлено у хворих на ОАГ, склавши в середньому 14,3±0,47 нг/мл, причому у 81% обстежених пацієнтів отримані результати суттєво перевищували показники, характерні для донорів, а також хворих на хронічний гепатит вірусної, змішаної, аутоімунної та алкогольної етіології (р<0,05), достигая 25 нг/мл.
Мал. 1. Концентрація ОБМ у хворих на ХГ залежно від етіології:

Мал. 2. Концентрація НСЕ у хворих на ХГ залежно від етіології:
1 – вірусний гепатит (HBV, HCV); 2 – аутоімунний гепатит; 3 – алкогольний гепатит;
4 – гепатит змішаної етіології; 5 - контроль
Висока концентрація в периферичній крові маркерів, що вивчаються, ушкодження нервової тканини, таких як ОБМ і НСЕ, виявлена нами при алкогольних ураженнях печінки, ймовірно, є проявом деміелінізуючих процесів, що нерідко спостерігаються при даній патології. Виявлені закономірності свідчать на користь того, що причинами розвитку атрофічних змін головного мозку та пошкоджень нервових волокон (маркерами яких є ОБМ та НСЕ), які часто зустрічаються в осіб, які зловживають алкоголем, є не тільки нейротоксичний ефект етанолу та його метаболітів, але й такі фактори, як дисфункція печінки, порушення харчування, а також дефіцит вітамінів групи В та нікотинової кислоти.
Як говорилося вище, основним етіологічним чинником, що призводить до виникнення хронічних гепатитів, є гепатотропна вірусна інфекція.
Показники концентрації ОБМ та НСЕ у сироватці крові хворих на ХГ залежно від виду гепатотропного вірусу (В та С) були близькі та достовірно не відрізнялися один від одного, а також від показників, отриманих у контролі (р>0,05). Також не було виявлено суттєвих відмінностей концентрацій досліджуваних маркерів деструкції нервової тканини у хворих на ХГС з 1 генотипом та генотипом «не-1» (2 та 3а). Отже, рівень у периферичній крові показників, що вивчаються нами, не залежить від типу вірусів.
Звертає на себе увагу, що показники концентрації ОБМ та НСЕ у хворих на ХГ вірусну та ХГ змішану етіологію (вірусну+алкогольну) суттєво не відрізняються один від одного, а також від результатів, отриманих у контролі (р>0,05). У той же час встановлено, що поєднання вірусного та алкогольного факторів надає більш істотний вплив на стан маркерів нейродеструкції, що вивчаються, ніж тільки при вірусній етіології. Так, якщо у хворих на змішану етіологію рівень ОБМ у 42% випадків перевищував показники, характерні для здорових осіб, то при хронічних вірусних гепатитах лише у 30%. Концентрація НСЕ відповідно у 39% випадків перевищувала показники, характерні для здорових осіб при змішаній етіології захворювання, а при вірусній лише у 31%. На наш погляд, це побічно свідчить про те, що висока концентрація маркерів, що вивчаються, ушкодження нервової тканини, виявлений у частини хворих на ХГ, більш характерна за наявності такого етіологічного фактора, як зловживання алкоголем.
Проведений у загальній групі хворих на ХГ кореляційний аналіз значень ОБМ та НСЕ показав відсутність достовірних зв'язків між цими показниками. У той же час у групі хворих з алкогольними ураженнями печінки було виявлено позитивну слабку силу кореляційний зв'язок між концентраціями ОБМ та НСЕ (r=0,45), що, на наш погляд, побічно свідчить про близькі механізми, що призводять до підвищення рівня цих маркерів ушкодження. нервової тканини при цій патології.
Виявлені закономірності дозволяють використовувати визначення рівня ОБМ та НСЕ у сироватці крові хворих на ХГ як додатковий маркер при діагностиці різних етіологічних форм хронічного гепатиту, насамперед алкогольної етіології, а також для виявлення наявності демієлінізуючих процесів при даній патології.
З огляду на те, що є етіологічні особливості характеру перебігу ЦП, темпів прогресування, розвитку ускладнень, було проведено вивчення концентрації ОБМ та НСЕ залежно від етіології захворювання. У 27 хворих (35%) був діагностований ЦП вірусної етіології, у 18 (23%) - алкогольний, у 22 (29%) в анамнезі зловживання алкоголем та перенесений вірусний гепатит одночасно (змішана етіологія), у 10 хворих (13%) був діагностовано первинний біліарний цироз печінки. Концентрації ОБМ та НСЕ у хворих на ЦП вірусної етіології склали 2,3±0,42 та 8,2±0,56 нг/мл, змішаної - 2,7±0,34 та 7,8±0,43 нг/мл, біліарної 3,2±0,39 та 8,3±0,39 нг/мл, алкогольної 3,4±0,3 та 8,9±044 нг/мл відповідно.
Середні значення концентрації НСЕ у групах хворих на ЦП вірусної, біліарної та алкогольної етіології достовірно (р<0,05) превышали показатели в контрольной группе. В то же время отсутствовали достоверные различия концентраций НСЕ в периферической крови в зависимости от этиологии ЦП. Результаты проведённого исследования свидетельствуют, что на стадии ЦП, в отличие от ХГ, концентрация данного маркера нейродеструкции в периферической крови не связана с этиологией заболевания.
Отже, на стадії сформованого ЦП причини, що зумовлюють підвищення рівня НСЕ у периферичній крові, дещо відрізняються від таких при гепатитах (ОАГ, ХГ). Ймовірно, провідну роль відіграє нейротоксична дія продуктів ендогенної інтоксикації, що циркулюють у крові при тяжкій дисфункції печінки, а не пряма дія етанолу та його метаболітів.
Крім того, що НСЕ насамперед відноситься до внутрішньоклітинних ензимів центральної нервової системи і вважається одним з найбільш специфічних показників її ушкодження, в той же час існує п'ять молекулярних форм ізоферментів НСЕ, які виявляються не тільки в нейронах, а й у нейроендокринних клітинах, скелетній мускулатурі, печінці. , еритроцитах та тромбоцитах, і коливання її загального рівня можуть бути пов'язані безпосередньо з тяжкою дисфункцією печінки та розвитком різних ускладнень, характерних для ЦП.
Результати, отримані при дослідженні рівня ОБМ у периферичній крові хворих на ЦП різної етіології, дещо відрізнялися.
Так, результати дослідження свідчать, що при ЦП біліарної (3,2±0,39 нг/мл) та алкогольної (3,4±0,3 нг/мл) етіології значення ОБМ достовірно підвищено порівняно з контрольною групою – 1,9 ±0,3 нг/мл та хворими на цироз печінки вірусної етіології - 2,3±0,42 нг/мл (р<0,05). При ЦП вирусной этиологии уровень ОБМ был наиболее низким, сопоставимым с показателями, полученными в контроле (р>0,05). При ЦП змішаної етіології (2,7±0,34 нг/мл) його рівень був дещо вищим, ніж при вірусній, і відповідно більшим, ніж у контролі, але достовірних відмінностей при зіставленні отриманих результатів не виявлено (р>0,05) . Незважаючи на достовірну різницю показників ОБМ крові у хворих на ЦП алкогольної етіології та ПБЦ у порівнянні з контролем, достовірної відмінності в рівні досліджуваного білка між цими групами хворих, що вивчаються, ми не виявили (р>0,05). Середні значення концентрації ОБМ у периферичній крові у хворих на цироз печінки змішаної та алкогольної етіології незначно відрізнялися один від одного: 2,7±0,34 та 3,4±0,3 нг/мл відповідно, достовірної відмінності виявлено не було (р>0 ,05). Отримані результати подано на рис. 3 та 4.

Мал. 3. Концентрація НСЕ у хворих на ЦП залежно від етіології: 1 - ЦП вірусної етіології (HBV, HCV); 2 – первинний біліарний цироз; 3 – ЦП алкогольної етіології; 4 – ЦП змішаної етіології; 5 - контроль

Мал. 4. Концентрація ОБМ у хворих на ЦП залежно від етіології: 1 - ЦП вірусної етіології (HBV, HCV); 2 – первинний біліарний цироз; 3 – ЦП алкогольної етіології; 4 – ЦП змішаної етіології; 5 - контроль
Таким чином, виявлені закономірності аналогічні результатам, отриманим у хворих на ХГ, у групі яких максимальна концентрація ОБМ плазми так само спостерігалася при алкогольній етіології захворювання.
Висновок. При алкогольних ураженнях печінки, як на стадії ХГ, так і ЦП, що сформувався, спостерігається значне підвищення концентрації ОБМ крові в порівнянні з вірусними ураженнями, що підтверджує наше припущення про те, що, крім нейротоксичної дії продуктів ендогенної інтоксикації, що циркулюють у крові при тяжких ураженнях печінки. Суттєву роль у процесах нейродеструкції та демієлінізації нервових волокон відіграє прямий ушкоджуючий ефект етанолу та його метаболітів.

Пропонуємо до вашої уваги журнали, що видаються у видавництві «Академія Природознавства»
Всі ліпіди, виявлені в мозку щури, присутні і в мієліну, тобто немає ліпідів, локалізованих виключно в немієлінових структурах (за винятком специфічного мітохондріального ліпіду дифосфатидилгліцерол). Вірне і зворотне - немає таких ліпідів мієліну, які не були б виявлені в інших субклітинних фракціях мозку.
Цереброзид – найбільш типовий компонент мієліну. За винятком раннього періоду розвитку організму, концентрація цереброзиду у мозку прямо пропорційна кількості у ньому мієліну. Тільки 1/5 загального вмісту галактоліпідів у мієліну зустрічається у сульфатованій формі. Цереброзиди та сульфатиди відіграють важливу роль у забезпеченні стабільності мієліну.
Для мієліну також характерний високий рівень його головних ліпідів - холестерину, загальних галактоліпідів і плазмалогену, що містить етаноламін. Встановлено, що до 70% холестерину мозку знаходиться у мієліну. Оскільки майже половина білої речовини мозку може складатися з мієліну, очевидно, що у мозку міститься найбільша кількість холестерину порівняно з іншими органами. Висока концентрація холестерину в мозку, особливо в мієліну, визначається основною функцією нейрональної тканини – генерувати та проводити нервові імпульси. Великий вміст холестерину в мієліну та своєрідність його структури призводять до зменшення іонного витоку через мембрану нейрона (внаслідок її високого опору).
Фосфатидилхолін також є суттєвою складовою мієліну, хоча сфінгомієлін міститься у відносно незначній кількості.
Ліпідний склад як сірої речовини, так і білої речовини мозку чітко відрізняється від такого у мієліну. Склад мієліну мозку всіх вивчених видів ссавців майже однаковий; мають місце лише незначні відмінності (наприклад, мієлін щура має менше сфінгомієліну, ніж мієлін бика чи людини). Існують деякі варіації і в залежності від локалізації мієліну, наприклад, мієлін, ізольований зі спинного мозку, має більш високе значення відношення ліпіду до білка, ніж мієлін з головного мозку.
До складу мієліну входять також поліфосфатидилінозити, з яких трифосфоінозитид становить від 4 до 6% загального фосфору мієліну, а дифосфоінозитид - від 1 до 1,5%. Мінорні компоненти мієліну включають принаймні три ефіри цереброзиду та два ліпіди на основі гліцерину; у складі мієліну також присутні деякі довголанцюгові алкани. Мієлін ссавців містить від 01 до 03% гангліозидів. У мієліну міститься більше моносиалоганглиозида вМ1 порівняно з тим, що виявляється у мембранах мозку. Мієлін багатьох організмів, у тому числі й людини, містить унікальний гангліозид сіалозилгалактозилцерамід ОМ4.
Ліпіди мієліну периферичної та центральної нервової системи якісно подібні, але між ними є кількісні відмінності. Мієлін ПНР містить менше цереброзидів і сульфатидів і значно більше сфінгомієліну, ніж мієлін ЦНС. Цікаво відзначити наявність гангліозиду ОМр характерного для мієліну ПНР деяких організмів. Відмінності у складі ліпідів мієліну центральної та периферичної нервової системи не такі істотні, як їх відмінності за білковим складом.
Білковий склад мієліну ЦНС більш простий, ніж інших мембран мозку, і представлений головним чином протеоліпідами та основними білками, які становлять 60-80% від загальної кількості. Глікопротеїни присутні у набагато менших кількостях. Мієлін центральної нервової системи містить унікальні білки.
Для мієліну ЦНС людини характерне кількісне превалювання двох білків: позитивно зарядженого катіонного білка мієліну (myelin basic protein, МВР) та протеоліпіду мієліну (myelin proteolipid protein, PLP). Ці білки - головні складові мієліну ЦНС всіх ссавців.
Мієліновий протеоліпід PLP (proteolipid protein), також відомий як білок Фолча, має здатність розчинятися в органічних розчинниках. Молекулярна маса PLP становить приблизно 30 кДа (Так – дальтон). Його амінокислотна послідовність є надзвичайно консервативною, молекула формує кілька доменів. Молекула PLP включає три жирні кислоти, як правило, пальмітинову, олеїнову та стеаринову, з'єднані з амінокислотними радикалами ефірним зв'язком.
Мієлін ЦНС містить дещо меншу кількість іншого протеоліпіду - DM-20, названого так за його молекулярною масою (20 кДа). І аналіз ДНК і з'ясування первинної структури показали, що DM-20 утворюється в результаті відщеплення 35 амінокислотних залишків від білка PLP. У процесі розвитку DM-20 з'являється раніше, ніж PLP (у деяких випадках навіть до появи мієліну); припускають, що на додаток до структурної ролі в освіті мієліну він може брати участь у диференціювання олігодендроцитів.
Всупереч уявленням про те, що PLP необхідний для формування компактного мультиламелярного мієліну, процес утворення мієліну у мишей, «нокаутованих» PLP/DM-20, відбувається лише з незначними відхиленнями. Однак у таких мишей зменшено тривалість життя та порушено загальну рухливість. Навпаки, природно мутації, що відбуваються в PLP, в тому числі його підвищена експресія (normal PLP over-expression), мають серйозні функціональні наслідки. Слід зазначити, що суттєві кількості білків PLP і DM-20 представлені в ЦНС, матрична РНК для PLP є і ПНС, і невелика кількість білка там синтезується, але не включається в мієлін.
Катіонний білок мієліну (МВР) привертає увагу дослідників внаслідок його антигенної природи - при введенні тварин він викликає аутоімунну реакцію, так званий експериментальний алергічний енцефаломієліт, який являє собою модель важкого нейродегенеративного захворювання - розсіяного склерозу.
Амінокислотна послідовність МВР у багатьох організмів високо консервативна. МВР розташований на цитоплазматичній стороні мієлінових мембран. Він має молекулярну масу 18,5 кДа та позбавлений ознак третинної структури. Цей основний білок виявляє мікрогетерогенність при електрофорез у лужних умовах. Більшість досліджених ссавців містили різні кількості ізоформ МБР, що мають суттєву загальну частину амінокислотної послідовності. Молекулярна маса МБР мишей та щурів – 14 кДа. МБР з малою молекулярною масою має такі ж амінокислотні послідовності на N- та С-термінальних частинах молекули, як і решті МБР, але відрізняється редукцією близько 40 амінокислотних залишків. Співвідношення цих основних білків змінюється у процесі розвитку: зрілі щури та миші мають більше МБР із молекулярною масою 14кДа, ніж МБР із молекулярною масою 18 кДа. Дві інші ізоформи МБР, які також виявляються в багатьох організмах, мають молекулярну масу 21,5 і 17 кДа, відповідно. Вони утворені приєднанням до основної структури поліпептидної послідовності масою близько 3 кДа.
При електрофоретичному розподілі білків мієліну виявляються білки з вищою молекулярною масою. Їхня кількість залежить від виду організму. Наприклад, миша та щур можуть містити таких білків до 30% від загальної кількості. Зміст цих білків також змінюється в залежності від віку тварини: чим вона молодша, тим менше в її мозку мієліну, але тим більше в ньому білків з більш високою молекулярною масою.
Фермент 2" 3"-циклічний нуклеотид З"-фосфодіестераза (CNP) становить кілька відсотків від загального вмісту мієлінового білка в клітинах ЦНС. Це набагато більше, ніж в інших типах клітин. Білок CNP - не головний компонент компактного мієліну, він сконцентрований лише в Білок локалізований в цитоплазмі, але частина його пов'язана з цитоскелетом мембрани - F-актином і тубуліном.Біологічна функція CNP може полягати в регулюванні структури цитоскелету для прискорення процесів росту.
Мієлінасоційований глікопротеїн (MAG) - мінорний у кількісному відношенні компонент очищеного мієліну, має молекулярну масу 100 кДа, міститься в ЦНС у невеликій кількості (менше 1% від загального білка). MAG має єдиний трансмембранний домен, який відокремлює сильноглікозильовану позаклітинну частину молекули, складену з п'яти подібних до доменів імуноглобуліну, від внутрішньоклітинного домену. Його повна структура подібна до білка адгезії нейрональної клітини (NCAM).
MAG не присутній у компактному, мультиламелярному мієліну, але знаходиться в періаксональних мембранах олігодендроцитів, що утворюють шари мієліну. Нагадаємо, що періаксональна мембрана олігодендроциту - найбільш близько розташована до плазматичної мембрани аксона, проте ці дві мембрани не зливаються, а розділені екстраклітинною щілиною. Подібна особливість локалізації MAG, а також те, що цей білок відноситься до імуноглобулінового суперсімейства, підтверджує участь його в процесах адгезії та передачі інформації (сигналінгу) між аксолеммою та мієлінутворюючими олігодендроцитами в процесі мієлінізації. Крім того, MAG - один із компонентів білої речовини ЦНС, що інгібує зростання нейритів у культурі тканини.
З інших глікопротеїнів білої речовини та мієліну слід відзначити мінорний мієлінолігодендроцитарний глікопротеїн (Myelin-oligodendrocytic glycoprotein, MOG). MOG є трансмембранним білком, що містить єдиний імуноглобулінподібний домен. На відміну від MAG, який розташований у внутрішніх шарах мієліну, MOG локалізований у поверхневих шарах, в силу чого може брати участь у передачі позаклітинної інформації до олігодендроциту.
Малі кількості характерних білків мембран можуть бути ідентифіковані внаслідок електрофорезу на поліакриламідному гелі (ПААГ) (наприклад, тубулін). Електрофорез високої роздільної здатності демонструє наявність інших незначних смуг білка; вони можуть бути пов'язані з присутністю ряду ферментів мієлінової оболонки.
Мієлін ПНР містить як деякі унікальні білки, так і кілька загальних з білками мієліну ЦНС білків.
Р0 - головний білок мієліну ПНР, має молекулярну масу 30 кДа, становить більше половини білків мієліну ПНР. Цікаво відзначити, що хоча він відрізняється від PLP по амінокислотній послідовності, шляхах посттрансляційної модифікації та структурі, проте обидва ці білки мають однаково важливе значення для формування структури мієліну ЦНС та ПНР.
Зміст МВР в мієлін ПНС становить 5-18% від загальної кількості білка, на відміну від ЦНС, де його частка досягає третини всього білка. Ті ж чотири форми білка МВР з молекулярними масами 21, 18,5, 17 і 14кДа, відповідно, виявлені в мієлін ЦНС, присутні і в ПНР. У дорослих гризунів МВР із молекулярною масою 14 кДа (за класифікацією периферійних мієлінових білків йому присвоєно назву «Рr») є найзначнішим компонентом усіх катіонних білків. У мієлін ПНС присутній і МВР з молекулярною масою 18 кДа (в цьому випадку він носить назву «білок Р1»). Слід зазначити, що важливість сімейства білків МВР не така велика для мієлінової структури ПНР, як для ЦНС.
Компактний мієлін ПНР містить глікопротеїн з молекулярною масою 22 кДа, названий периферійним мієліновим білком 22 (РМР-22), частка якого становить менше 5% від загального вмісту білків. РМР-22 має чотири трансмембранні домени і один глікозильований домен. Цей білок не має значної структурної ролі. Однак аномалії гена рMR-22 відповідальні за деякі спадкові невропатології людини.
Декілька десятиліть тому вважалося, що мієлін створює інертну оболонку, яка не виконує жодних біохімічних функцій. Однак пізніше в мієліну було виявлено велику кількість ферментів, що залучаються до синтезу та метаболізму компонентів мієліну. Ряд ферментів, присутніх у мієліну, включається в метаболізм фосфоінозитідів: фосфатидилінозитолкіназа, дифосфатидилінозитолкіназа, відповідні фосфатази та дигліцеридкінази. Ці ферменти становлять інтерес внаслідок високої концентрації в мієлін поліфосфоінозітідів та їх швидкого обміну. Є свідчення присутності в мієліну мускаринових холінергічних рецепторів, G-білків, фосфоліпаз С та Е, протеїнкінази С.
У мієліну ПНЗ виявлено Nа/К-АТФазу, що здійснює транспорт одновалентних катіонів, а також 6"-нуклеотидазу. Наявність цих ферментів дозволяє припустити, що мієлін може брати активну участь в аксональному транспорті.
| Статті на тему: | |
|
Хто призначає дослідження
Захворювання, одним з основних проявів яких є... Миш'як – небезпечна, але необхідна речовина Розповсюдження миш'яку в природі
Миш'як МИШ'ЯК -а; м. 1. Хімічний елемент (Аs) - тверде... Гафнієва чудо-бомба та сувора реальність Графітова бомба як працює
Протягом двох місяців авіація НАТО наносила так звані... | |




